
Via het Linkedin account van Mario Ortiz Martinez vond ik de volgende productie waarin wordt uitgelegd dat het Pfizer vaccin het DNA van de lever aantast. Voorts wordt de Neurenberg Code aangehaald, waarin duidelijk is weergegeven dat het iedereen vrij moet staan om wel dan niet te kiezen voor een vaccinatie en gezien de tekst zit daar geen woord Spaans bij (overigens heeft de EU eerder laten weten tegen verpolichte vaccinatie te zijn, al is die mening intussen losgelaten door hare kwaadaardigheid von der Leyen, althans als ik me niet vergis):
"1. The voluntary consent of the human subject is absolutely essential. This means that the person involved should have legal capacity to give consent; should be so situated as to be able to exercise free power of choice, without the intervention of any element of force, fraud, deceit, duress, over-reaching, or other ulterior form of constraint or coercion."
Deze tekst kom je hieronder nogmaals tegen en gezien de volledige tekst is het m.i. onverantwoord om je te laten vaccineren met het Pfizer vaccin. Maar lees de tekst en vorm je eigen oordeel. (zie alvorens tot vaccinatie over te gaan ook de berichten onder de door mij toegevoegde links en ook daarbij: vorm je eigen mening)
(On the top right hand side of this page you can choose for a translation in the language of your choice: choose 'Engels' [english] so you can recognise your own language [the Google translation is first in dutch, a language most people don't understand, while on the other hand most people recognise there language translated in english])
(als je het Engels niet machtig bent, kopieer dan de Engelse tekst en plak die in deze vertaalapp, de app werkt snel en de vertaling is van een redelijk goede kwaliteit. Kopieer het artikel en plaats het links in de app, je ziet rechts Italiaans staan als je daar op klikt kan je kiezen voor Nederlands)
Hier de productie van MDPI:
Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line

 , 1, 1, 1, 1, 2 and 1,*
, 1, 1, 1, 1, 2 and 1,*
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. REAL-TIME RT-QPCR

| Target | Sequence |
|---|---|
| ACTB forward | CCTCGCCTTTGCCGATCC |
| ACTB reverse | GGATCTTCATGAGGTAGTCAGTC |
| GAPDH forward | CTCTGCTCCTCCTGTTCGAC |
| GAPDH reverse | TTAAAAGCAGCCCTGGTGAC |
| LINE-1 forward | TAACCAATACAGAGAAGTGC |
| LINE-1 reverse | GATAATATCCTGCAGAGTGT |
| BNT162b2 forward | CGAGGTGGCCAAGAATCTGA |
| BNT162b2 reverse | TAGGCTAAGCGTTTTGAGCTG |
2.3. Immunofluorescence Staining and Confocal Imaging
2.4. Genomic DNA Purification, PCR Amplification, Agarose Gel Purification, and Sanger Sequencing
Statistics
2.5. Ethical Statements
3. Results
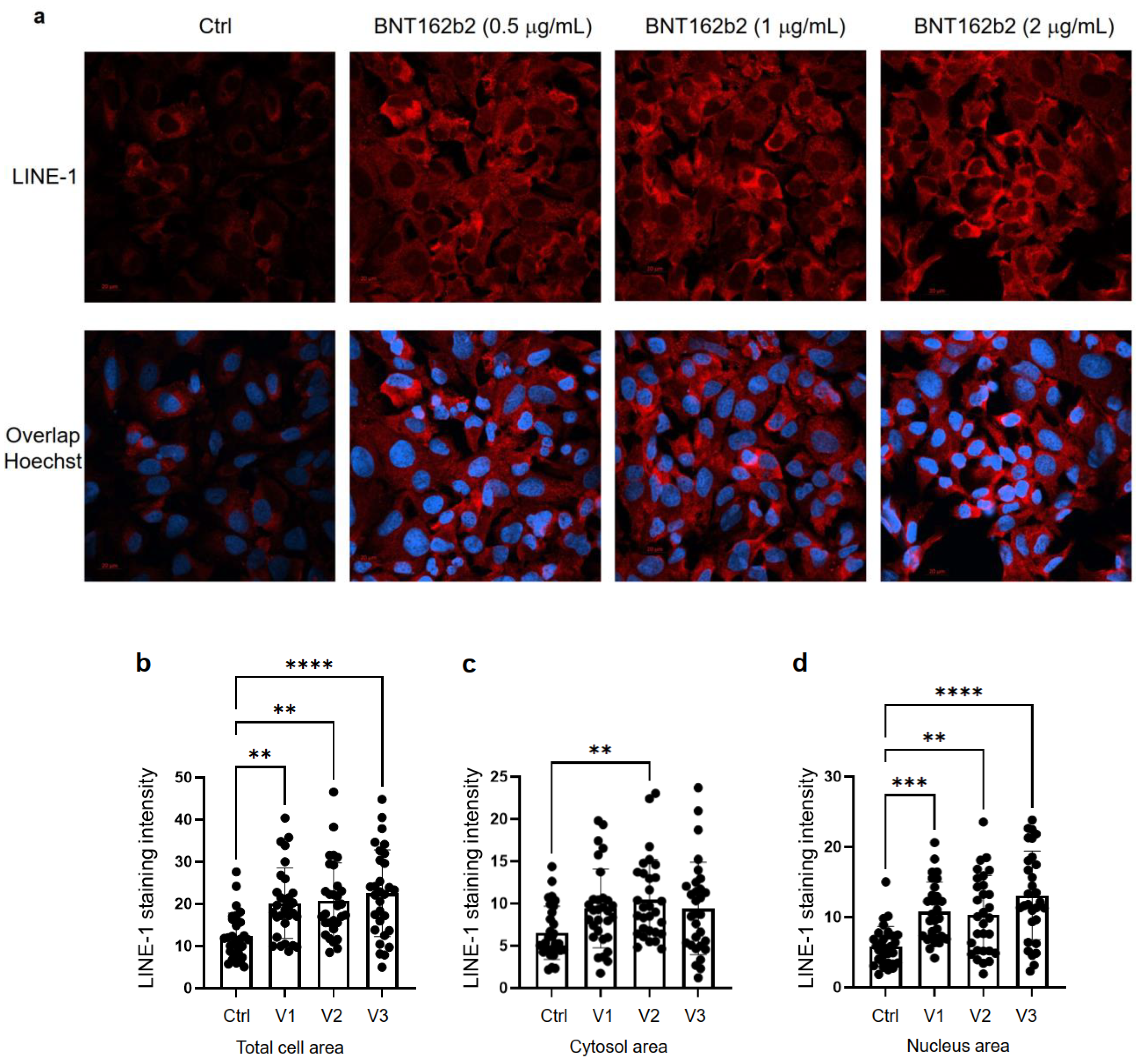
3.1. BNT162b2 Enters Human Liver Cell Line Huh7 Cells at High Efficiency
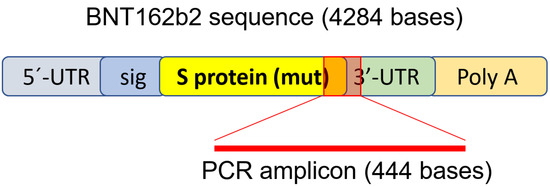
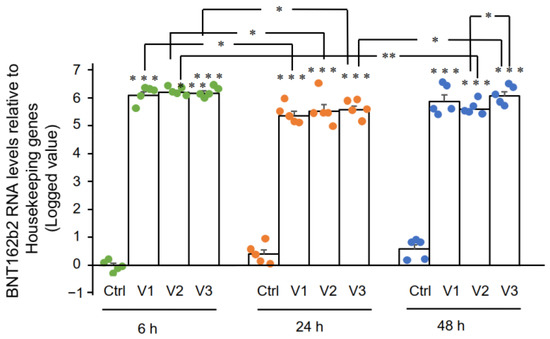
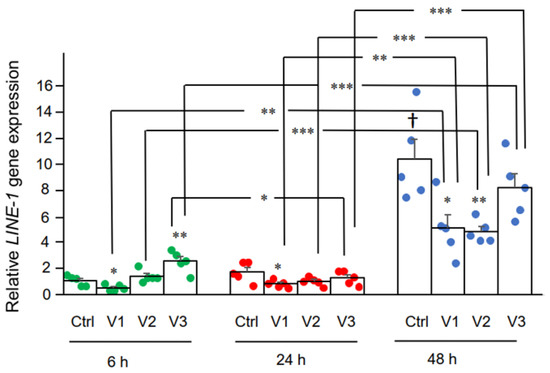
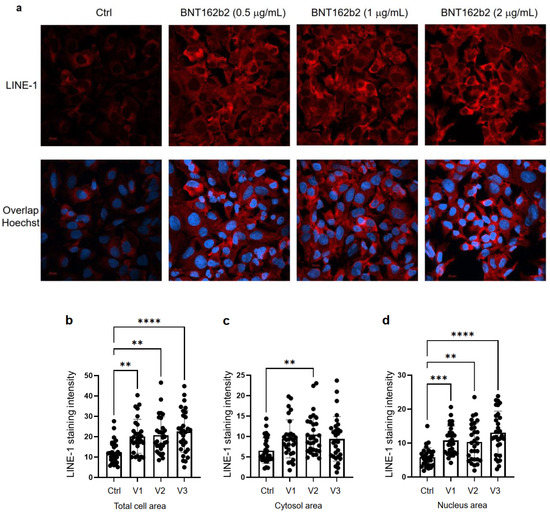
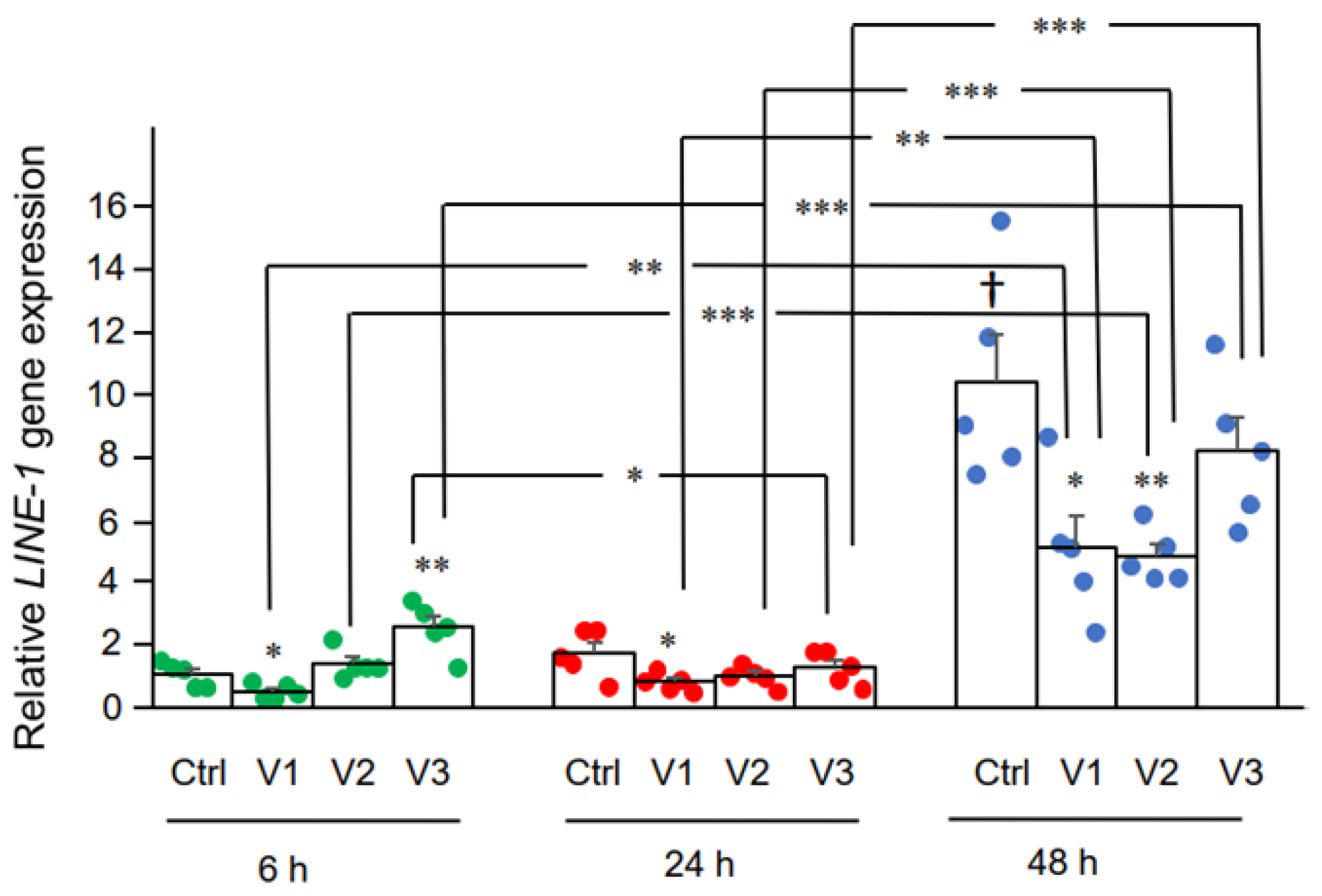
3.2. Effect of BNT162b2 on Human Endogenous Reverse Transcriptase Long Interspersed Nuclear Element-1 (LINE-1)
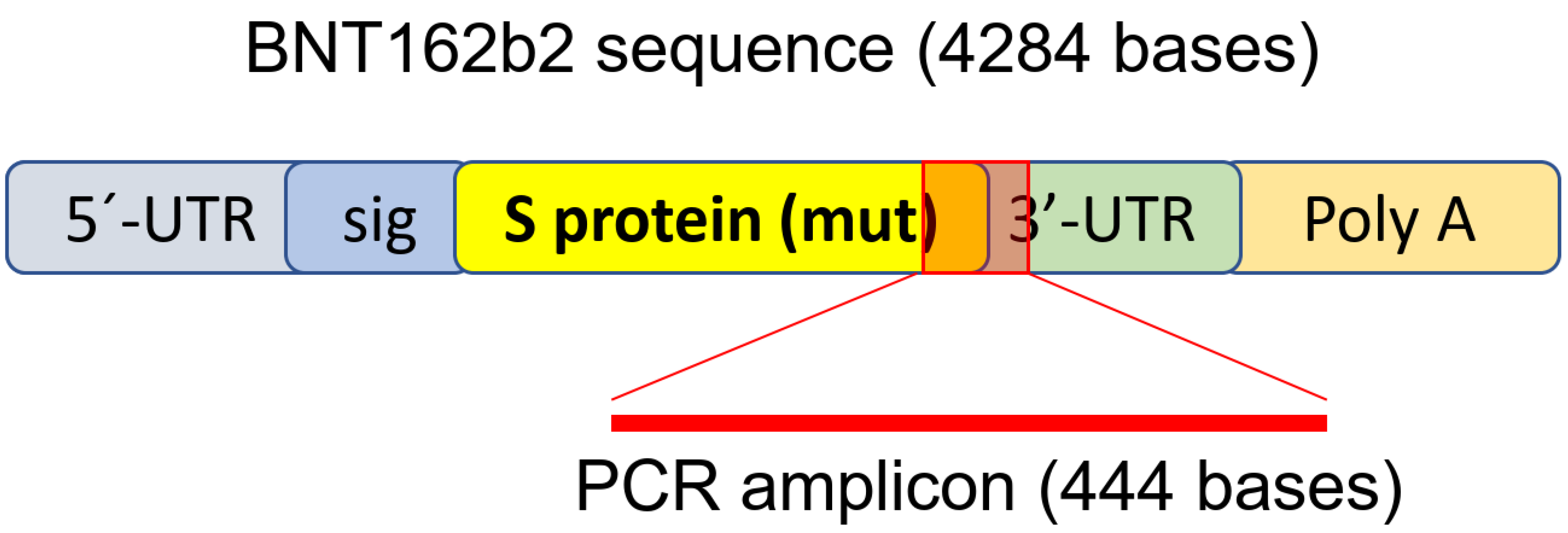
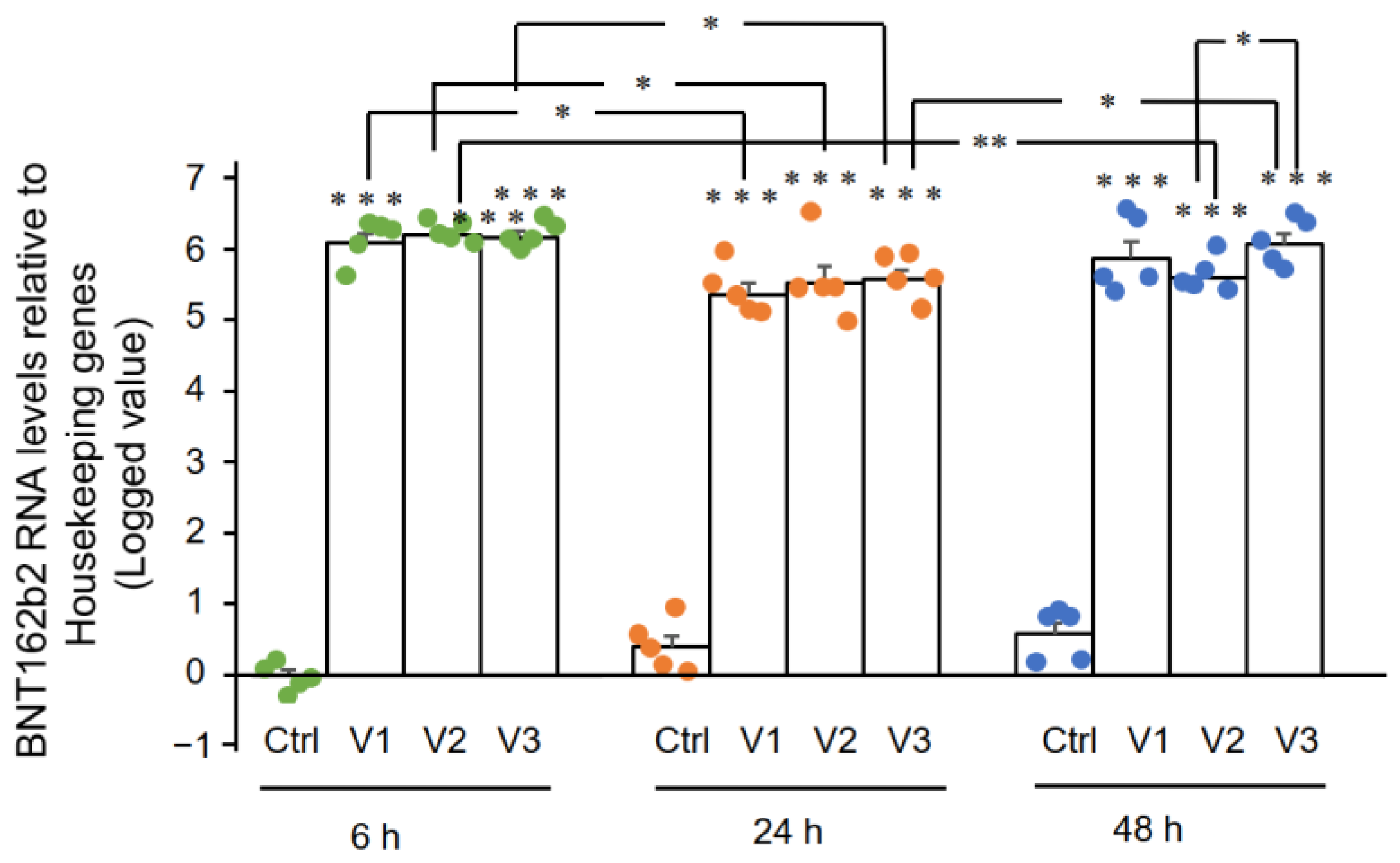
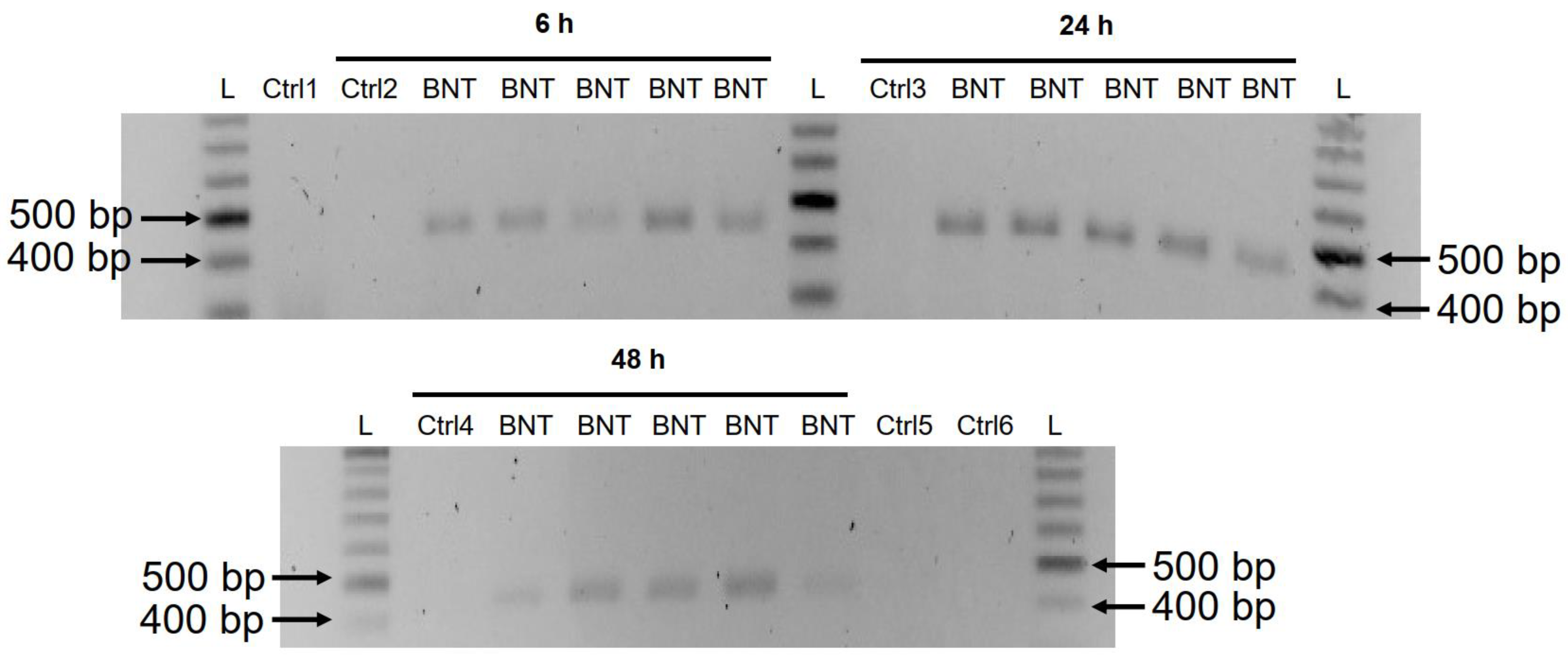
3.3. Detection of Reverse Transcribed BNT162b2 DNA in Huh7 Cells
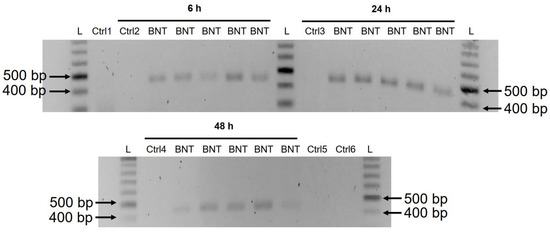
| CGAGGTGGCCAAGAATCTGAACGAGAGCCTGATCGACCTGCAAGAACTGGGGAAGT ACGAGCAGTACATCAAGTGGCCCTGGTACATCTGGCTGGGCTTTATCGCCGGACTGATTG CCATCGTGATGGTCACAATCATGCTGTGTTGCATGACCAGCTGCTGTAGCTGCCTGAAGG GCTGTTGTAGCTGTGGCAGCTGCTGCAAGTTCGACGAGGACGATTCTGAGCCCGTGCTGA |
| AGGGCGTGAAACTGCACTACACATGATGACTCGAGCTGGTACTGCATGCACGCAATGCTA GCTGCCCCTTTCCCGTCCTGGGTACCCCGAGTCTCCCCCGACCTCGGGTCCCAGGTATGC TCCCACCTCCACCTGCCCCACTCACCACCTCTGCTAGTTCCAGACACCTCCCAAGCACGC AGCAATGCAGCTCAAAACGCTTAGCCTA |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 22 February 2022).
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Hall, J.A.; Zaidi, A.; Andrews, N.J.; Dunbar, J.K.; Dabrera, G. Effect of Vaccination on Household Transmission of SARS-CoV-2 in England. N. Engl. J. Med. 2021, 385, 759–760. [Google Scholar] [CrossRef]
- Butt, A.A.; Omer, S.B.; Yan, P.; Shaikh, O.S.; Mayr, F.B. SARS-CoV-2 Vaccine Effectiveness in a High-Risk National Population in a Real-World Setting. Ann. Intern. Med. 2021, 174, 1404–1408. [Google Scholar] [CrossRef]
- Dagan, N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernan, M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef]
- Rossman, H.; Shilo, S.; Meir, T.; Gorfine, M.; Shalit, U.; Segal, E. COVID-19 dynamics after a national immunization program in Israel. Nat. Med. 2021, 27, 1055–1061. [Google Scholar] [CrossRef]
- Fan, B.E.; Shen, J.Y.; Lim, X.R.; Tu, T.M.; Chang, C.C.R.; Khin, H.S.W.; Koh, J.S.; Rao, J.P.; Lau, S.L.; Tan, G.B.; et al. Cerebral venous thrombosis post BNT162b2 mRNA SARS-CoV-2 vaccination: A black swan event. Am. J. Hematol. 2021, 96, E357–E361. [Google Scholar] [CrossRef]
- Larson, K.F.; Ammirati, E.; Adler, E.D.; Cooper, L.T., Jr.; Hong, K.N.; Saponara, G.; Couri, D.; Cereda, A.; Procopio, A.; Cavalotti, C.; et al. Myocarditis After BNT162b2 and mRNA-1273 Vaccination. Circulation 2021, 144, 506–508. [Google Scholar] [CrossRef]
- Menni, C.; Klaser, K.; May, A.; Polidori, L.; Capdevila, J.; Louca, P.; Sudre, C.H.; Nguyen, L.H.; Drew, D.A.; Merino, J.; et al. Vaccine side-effects and SARS-CoV-2 infection after vaccination in users of the COVID Symptom Study app in the UK: A prospective observational study. Lancet Infect. Dis. 2021, 21, 939–949. [Google Scholar] [CrossRef]
- Hansen, T.; Titze, U.; Kulamadayil-Heidenreich, N.S.A.; Glombitza, S.; Tebbe, J.J.; Rocken, C.; Schulz, B.; Weise, M.; Wilkens, L. First case of postmortem study in a patient vaccinated against SARS-CoV-2. Int. J. Infect. Dis. 2021, 107, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Kadali, R.A.K.; Janagama, R.; Peruru, S.; Malayala, S.V. Side effects of BNT162b2 mRNA COVID-19 vaccine: A randomized, cross-sectional study with detailed self-reported symptoms from healthcare workers. Int. J. Infect. Dis. 2021, 106, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Parkash, O.; Sharko, A.; Farooqi, A.; Ying, G.W.; Sura, P. Acute Pancreatitis: A Possible Side Effect of COVID-19 Vaccine. Cureus 2021, 13, e14741. [Google Scholar] [CrossRef] [PubMed]
- Mazzatenta, C.; Piccolo, V.; Pace, G.; Romano, I.; Argenziano, G.; Bassi, A. Purpuric lesions on the eyelids developed after BNT162b2 mRNA COVID-19 vaccine: Another piece of SARS-CoV-2 skin puzzle? J. Eur. Acad. Dermatol. Venereol. 2021, 35, e543–e545. [Google Scholar] [CrossRef]
- Lee, E.J.; Cines, D.B.; Gernsheimer, T.; Kessler, C.; Michel, M.; Tarantino, M.D.; Semple, J.W.; Arnold, D.M.; Godeau, B.; Lambert, M.P.; et al. Thrombocytopenia following Pfizer and Moderna SARS-CoV-2 vaccination. Am. J. Hematol. 2021, 96, 534–537. [Google Scholar] [CrossRef]
- Ishay, Y.; Kenig, A.; Tsemach-Toren, T.; Amer, R.; Rubin, L.; Hershkovitz, Y.; Kharouf, F. Autoimmune phenomena following SARS-CoV-2 vaccination. Int. Immunopharmacol. 2021, 99, 107970. [Google Scholar] [CrossRef]
- Das, B.B.; Kohli, U.; Ramachandran, P.; Nguyen, H.H.; Greil, G.; Hussain, T.; Tandon, A.; Kane, C.; Avula, S.; Duru, C.; et al. Myopericarditis following mRNA COVID-19 Vaccination in Adolescents 12 through 18 Years of Age. J. Pediatr. 2021, 238, 26–32.e1. [Google Scholar] [CrossRef]
- McLaurin-Jiang, S.; Garner, C.D.; Krutsch, K.; Hale, T.W. Maternal and Child Symptoms Following COVID-19 Vaccination Among Breastfeeding Mothers. Breastfeed. Med. 2021, 16, 702–709. [Google Scholar] [CrossRef]
- Barda, N.; Dagan, N.; Ben-Shlomo, Y.; Kepten, E.; Waxman, J.; Ohana, R.; Hernan, M.A.; Lipsitch, M.; Kohane, I.; Netzer, D.; et al. Safety of the BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Setting. N. Engl. J. Med. 2021, 385, 1078–1090. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, S.; Warkentin, T.E.; Greinacher, A. Thrombotic Thrombocytopenia after ChAdOx1 nCoV-19 Vaccination. Reply. N. Engl. J. Med. 2021, 385, e11. [Google Scholar] [CrossRef] [PubMed]
- Doroftei, B.; Ciobica, A.; Ilie, O.D.; Maftei, R.; Ilea, C. Mini-Review Discussing the Reliability and Efficiency of COVID-19 Vaccines. Diagnostics 2021, 11, 579. [Google Scholar] [CrossRef]
- Zhang, L.; Richards, A.; Barrasa, M.I.; Hughes, S.H.; Young, R.A.; Jaenisch, R. Reverse-transcribed SARS-CoV-2 RNA can integrate into the genome of cultured human cells and can be expressed in patient-derived tissues. Proc. Natl. Acad. Sci. USA 2021, 118, e2105968118. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.ema.europa.eu/en/documents/assessment-report/comirnaty-epar-public-assessment-report_en.pdf (accessed on 24 February 2022).
- Tanaka, H.; Takata, N.; Sakurai, Y.; Yoshida, T.; Inoue, T.; Tamagawa, S.; Nakai, Y.; Tange, K.; Yoshioka, H.; Maeki, M.; et al. Delivery of Oligonucleotides Using a Self-Degradable Lipid-Like Material. Pharmaceutics 2021, 13, 544. [Google Scholar] [CrossRef]
- Sedic, M.; Senn, J.J.; Lynn, A.; Laska, M.; Smith, M.; Platz, S.J.; Bolen, J.; Hoge, S.; Bulychev, A.; Jacquinet, E.; et al. Safety Evaluation of Lipid Nanoparticle-Formulated Modified mRNA in the Sprague-Dawley Rat and Cynomolgus Monkey. Vet. Pathol. 2018, 55, 341–354. [Google Scholar] [CrossRef]
- Sato, Y.; Matsui, H.; Yamamoto, N.; Sato, R.; Munakata, T.; Kohara, M.; Harashima, H. Highly specific delivery of siRNA to hepatocytes circumvents endothelial cell-mediated lipid nanoparticle-associated toxicity leading to the safe and efficacious decrease in the hepatitis B virus. J. Control. Release 2017, 266, 216–225. [Google Scholar] [CrossRef]
- Heidel, J.D.; Yu, Z.; Liu, J.Y.; Rele, S.M.; Liang, Y.; Zeidan, R.K.; Kornbrust, D.J.; Davis, M.E. Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc. Natl. Acad. Sci. USA 2007, 104, 5715–5721. [Google Scholar] [CrossRef]
- Available online: https://www.cvdvaccine-us.com/ (accessed on 24 February 2022).
- Available online: http://bridgeslab.sph.umich.edu/protocols/index.php/Preparation_of_Tail_Samples_(for_Genotyping) (accessed on 24 February 2022).
- Gallud, A.; Munson, M.J.; Liu, K.; Idstrom, A.; Barriga, H.M.; Tabaei, S.; Aliakbarinodehi, N.; Ojansivu, M.; Lubart, Q.; Doutch, J.J.; et al. Time evolution of PEG-shedding and serum protein coronation determines the cell uptake kinetics and delivery of lipid nanoparticle. bioRxiv 2021. [Google Scholar] [CrossRef]
- World Health Organization Messenger RNA Encoding the Full-Length SARS-CoV-2 Spike Glycoprotein. 2020. Available online: https://web.archive.org/web/20210105162941/https://mednet-communities.net/inn/db/media/docs/11889.doc (accessed on 24 February 2022).
- Mita, P.; Wudzinska, A.; Sun, X.; Andrade, J.; Nayak, S.; Kahler, D.J.; Badri, S.; LaCava, J.; Ueberheide, B.; Yun, C.Y.; et al. LINE-1 protein localization and functional dynamics during the cell cycle. Elife 2018, 7, e30058. [Google Scholar] [CrossRef]
- Sato, Y.; Kinami, Y.; Hashiba, K.; Harashima, H. Different kinetics for the hepatic uptake of lipid nanoparticles between the apolipoprotein E/low density lipoprotein receptor and the N-acetyl-d-galactosamine/asialoglycoprotein receptor pathway. J. Control. Release 2020, 322, 217–226. [Google Scholar] [CrossRef]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Guler, A.; et al. BNT162b vaccines protect rhesus macaques from SARS-CoV-2. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Bahl, K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.; Laska, M.E.; Smith, M.; Almarsson, O.; Thompson, J.; et al. Preclinical and Clinical Demonstration of Immunogenicity by mRNA Vaccines against H10N8 and H7N9 Influenza Viruses. Mol. Ther. 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Bril, F.; Al Diffalha, S.; Dean, M.; Fettig, D.M. Autoimmune hepatitis developing after coronavirus disease 2019 (COVID-19) vaccine: Causality or casualty? J. Hepatol. 2021, 75, 222–224. [Google Scholar] [CrossRef]
- Kazazian, H.H., Jr.; Moran, J.V. Mobile DNA in Health and Disease. N. Engl. J. Med. 2017, 377, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.M.; Fan, H. The Discovery of Reverse Transcriptase. Annu. Rev. Virol. 2016, 3, 29–51. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Ostertag, E.M.; Goodier, J.L.; Zhang, Y.; Kazazian, H.H., Jr. SVA elements are nonautonomous retrotransposons that cause disease in humans. Am. J. Hum. Genet. 2003, 73, 1444–1451. [Google Scholar] [CrossRef]
- Hancks, D.C.; Kazazian, H.H., Jr. Active human retrotransposons: Variation and disease. Curr. Opin. Genet. Dev. 2012, 22, 191–203. [Google Scholar] [CrossRef]
- Jones, R.B.; Song, H.; Xu, Y.; Garrison, K.E.; Buzdin, A.A.; Anwar, N.; Hunter, D.V.; Mujib, S.; Mihajlovic, V.; Martin, E.; et al. LINE-1 retrotransposable element DNA accumulates in HIV-1-infected cells. J. Virol. 2013, 87, 13307–13320. [Google Scholar] [CrossRef]
- Macchietto, M.G.; Langlois, R.A.; Shen, S.S. Virus-induced transposable element expression up-regulation in human and mouse host cells. Life Sci. Alliance 2020, 3, e201900536. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, X.Z.; He, X.; Zhou, L.Q. Exogenous Coronavirus Interacts With Endogenous Retrotransposon in Human Cells. Front. Cell Infect. Microbiol. 2021, 11, 609160. [Google Scholar] [CrossRef] [PubMed]
- Belancio, V.P.; Roy-Engel, A.M.; Deininger, P. The impact of multiple splice sites in human L1 elements. Gene 2008, 411, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Taylor, M.S.; O’Donnell, K.A.; Boeke, J.D. Poly(A) binding protein C1 is essential for efficient L1 retrotransposition and affects L1 RNP formation. Mol. Cell Biol. 2012, 32, 4323–4336. [Google Scholar] [CrossRef]
- Servant, G.; Streva, V.A.; Derbes, R.S.; Wijetunge, M.I.; Neeland, M.; White, T.B.; Belancio, V.P.; Roy-Engel, A.M.; Deininger, P.L. The Nucleotide Excision Repair Pathway Limits L1 Retrotransposition. Genetics 2017, 205, 139–153. [Google Scholar] [CrossRef]
- Guo, H.; Chitiprolu, M.; Gagnon, D.; Meng, L.; Perez-Iratxeta, C.; Lagace, D.; Gibbings, D. Autophagy supports genomic stability by degrading retrotransposon RNA. Nat. Commun. 2014, 5, 5276. [Google Scholar] [CrossRef]
- Xie, Y.; Mates, L.; Ivics, Z.; Izsvak, Z.; Martin, S.L.; An, W. Cell division promotes efficient retrotransposition in a stable L1 reporter cell line. Mob. DNA 2013, 4, 10. [Google Scholar] [CrossRef]
- Shi, X.; Seluanov, A.; Gorbunova, V. Cell divisions are required for L1 retrotransposition. Mol. Cell Biol. 2007, 27, 1264–1270. [Google Scholar] [CrossRef]
- Goff, S.P. Host factors exploited by retroviruses. Nat. Rev. Microbiol 2007, 5, 253–263. [Google Scholar] [CrossRef]
- Suzuki, Y.; Craigie, R. The road to chromatin—Nuclear entry of retroviruses. Nat. Rev. Microbiol. 2007, 5, 187–196. [Google Scholar] [CrossRef]
- Shi, J.; Wang, X.; Lyu, L.; Jiang, H.; Zhu, H.J. Comparison of protein expression between human livers and the hepatic cell lines HepG2, Hep3B, and Huh7 using SWATH and MRM-HR proteomics: Focusing on drug-metabolizing enzymes. Drug Metab. Pharmacokinet. 2018, 33, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Kubo, S.; Seleme, M.C.; Soifer, H.S.; Perez, J.L.; Moran, J.V.; Kazazian, H.H., Jr.; Kasahara, N. L1 retrotransposition in nondividing and primary human somatic cells. Proc. Natl. Acad. Sci. USA 2006, 103, 8036–8041. [Google Scholar] [CrossRef] [PubMed]
- Macia, A.; Widmann, T.J.; Heras, S.R.; Ayllon, V.; Sanchez, L.; Benkaddour-Boumzaouad, M.; Munoz-Lopez, M.; Rubio, A.; Amador-Cubero, S.; Blanco-Jimenez, E.; et al. Engineered LINE-1 retrotransposition in nondividing human neurons. Genome Res. 2017, 27, 335–348. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
==========================================
Meer over het farmaceutische maffia lid Pfizer: 'Pfizer CEO: Israëliërs zijn proefkonijn voor testen van vaccin' (!!!!) De CEO van Pfizer die zonder blikken of blozen aangeeft waar het om gaat als men gevaccineerd is....... Israël
waar intussen velen al spijt hebben zich te hebben laten
vaccineren..... Intussen heeft Pfizer afgezien van een contract met
India, daar het land extra garanties wilde over de veiligheid en daar
zag Pfizer geen brood in...... India een land met een bevolking van 1,3
miljard mensen, ofwel daar was genoeg winst te behalen voor Pfizer; moet
je nagaan..... Er is nog een groot land dat extra veiligheid eiste,
waarop Pfizer ook de stekker uit die onderhandelingen trok..... Als je
hier naar zoekt op het net, maakt niet uit op wat voor manier, vind je
hier niets over terug, tekenend........
'Pfizer CEO calls people who spread vaccine misinformation 'criminals'' Een lid van de farmaceutische maffia die anderen aanwijst als misdadigers..... ha! ha! ha!
'COVID-19: FDA vraagt
om volledig rapport over toelating van het Pfizer vaccin tot 2076 geheim
te houden >> klokkenluider ontslagen' (!!!!) Ook die zaak
wekt 'veel vertrouwen' bij de bevolking, vertrouwen in het vaccin, de
Biden administratie en overheidsinstanties als de FDA....... Zoveel
vertrouwen als het volgende:
'COVID-19-vaccins: Pfizer maakt misbruik van wanhopige regeringen'
'COVID-19: Pfizer adviseert derde vaccinatie, omzet van dit bedrijf op deze vaccins in 2021 al meer dan 20 miljard dollar.......' (een bericht van 9 juli jl. 2021.....)
'Pfizer Coronavirus vaccin: wat je niet wordt verteld over de vaccins tegen COVID-19' In augustus 2020 deed Pfizer CEO Bourla 62% van z'n aandelen Pfizer van de hand, blijkbaar heeft hij er geen vertrouwen in dat de beloften van overheden dat het bedrijf niet vervolgd zal worden voor ernstige bijwerkingen ook zullen worden geëerbiedigd door rechters mocht men het bedrijf aanklagen voor doden als gevolg van vaccinatie dan wel voor ernstige (chronische) bijwerkingen......
'COVID-maatregelen: een open brief aan Ernst Kuipers'
'Aartsbisschop Carlo Maria Figanò: “Twee jaar lang zijn we nu getuige van een globale staatsgreep”'
(door de Coronamaatregelen) Overigens onbegrijpelijk dat deze
aartsbisschop zich in de bijgevoegde video alleen tot katholieken
richt......
'COVID-19: onderzoeken bevestigen dat natuurlijke verkregen immuniteit meer dan voldoende is' (!!!!) Op 15 februari 2022, werd bekendgemaakt dat natuurlijke
immuniteit veel langer standhoudt dan die middels een vaccin verkregen
en dat niet misselijk: volgens onderzoekers in Italië zelfs 18 maanden!! >> 'Natural Immunity Lasts for at Least 18 Months: Study' En wat
heeft Nederland gedaan?Juist, die periode van natuurlijke immuniteit
teruggebracht naar een half jaar!! Gelukkig zijn we voorlopig af van die
belachelijke beslissing!!
'Onverdeeld Open: teken het manifest om het Coronatoegangsbewijs onmiddellijk af te schaffen'
'Hans Teeuwen: "Geen Coronapas"'
'Intrekken uitzendlicentie door Rusland voor Deutsche Welle 'is een klap in het gezicht van het Duitse volk'' RT Deutsch dat werd platgelegd in Duitsland wordt o.a. verweten complottheorieën over COVID te brengen, dit door het uitzenden van een gesprek met een paar COVID-vaccinatieweigeraars....... (te zot voor woorden!!) Oh, en Deutsche Welle is een Duitse staatsomroep die westerse propaganda uitzond in Rusland, iets waarover men niet lult in het westen en als men er al over spreeekt wordt er onomwonden gezegd dat dit soort westerse platforms onafhankelijk zijn..... ha! ha! ha! ha! ha! ha! ha!
'EU medicijnregelgever EMA en de WHO roepen op te stoppen met COVID boosters vanwege aantasting immuunsysteem en falen bij bestrijding Coronavirus' (!!!!) zie wat dit betreft ook:
'COVID-19 vaccin zou het immuunsysteem aantasten.......' (!!!!) Benieuwd ook of de vandaag (het is terwijl ik dit schrijf 11 januari 2022 /////) overleden David Sassoli, voorzitter van het EU parlement, 3 dubbel gevaccineerd was, daar ook hij een afwijking had aan het immuunsysteem......
'COVID-19: IC verpleegkundige OLVG slaakt noodkreet'
Hier nog wat meer links naar 'oudere berichten': 'COVID-19: Gezondheidsraad houdt zich niet aan de regels van de Raad van Europa: discriminatie van niet-gevaccineerden is verboden' (!!!!)
'Besmette gevaccineerden houden 'gewoon' toegang tot openbare gelegenheden'
'Coronavirus: Diederik Gommers (IC arts) zou het mooi vinden dat mensen worden ingeënt met het AZ vaccin nadat ze tekenden voor het risico dat ze lopen.....' Dan te bedenken dat zelfs de farmaceuten hun eigen vaccin niet vertrouwen, zie wat dat betreft:
'Coronavaccinatie ook als je weet dat de fabrikant ernstige bijwerkingen niet uitsluit?' En zie:
'Coronavirus: paniek over de Deltavariant is onterecht en zwaar overdreven' (!!!!) En zie de links in dat bericht naar artikelen over de PCR test en de bezuinigingen op de gezondheidszorg >> die bezuinigingen zijn de reden voor de Coronamaatregelen, die werden immers genomen daar er te weinig ziekenhuisbedden, IC bedden en verpleegkundigen waren. Zoals ook in het bericht hierboven al gemeld: het aantal verpleegkundigen is als de bedden sterk teruggelopen door wanbeleid van de afbraakkabinetten Rutte 1, 2 en 3..... Zo waren er bij aantreden van Rutte 1 nog 2.200 IC bedden, dat was bij aanvang van de Coronacrisis teruggebracht naar 1.100 bedden..... Dus als je sterk bent benadeeld door de Coronamaatregelen, weet je wie hiervoor verantwoordelijk zijn!! Zie wat betreft dat bericht ook: 'Coronavirus: artsen willen via rechtszaak tegen de staat COVID-19 van A-lijst krijgen, daar het een griepvirus zou zijn 'De video die oorspronkelijk in het artikel was te zien is gecensureerd, tja je wilt natuurlijk niet dat de bevolking begrijpt hoezeer men werd en wordt belazerd......
'Met COVID-19 besmette zorgmedewerkers mogen ondanks symptomen gewoon doorwerken.......'
'Coronavirus: Fort Detrick eindelijk als verdachte van oorsprong aangemerkt'
'Vaccinatie tegen Spaanse Griep: miljoenen doden'
'COVID vaccinmakers: hoe betrouwbaar zijn deze farmaceuten' (en zie de links in dat bericht)
'COVID-19: gezonde mensen maken elkaar niet ziek'
'Coronabeslommeringen: ook na vaccinatie kan je het virus doorgeven, plus de zoveelste blunder van CDA minister de Jonge'
Het na dit bericht gebrachte nieuws dat je niet besmettelijk bent na
vaccinatie, is achteraf gebeleken grote onzin te zijn en zou gebaseerd
zijn op een
foute interpretatie van een bericht uit de VS, lijkt me ook logisch daar
dr Fauci eerder al meldde dat gevaccineerden nog lang besmettelijk
blijven en zich aan de Coronamaatregelen dienen te houden, terwijl de
voormaligfe minister van Volksgezodnheid, CDA plork fde Jonge vorig jaar
zelf toegaf dat gevaccineerden niet veilig zijn...... (zie de
afbeelding met deze zwaar disfunctionerende kwezel in het blok met Pfizer links hierboven)
'Coronavirus, PCR test: leugens en onzin' (een artikel van Mario Ortiz Buijsse)
'Met COVID-19 besmette zorgmedewerkers mogen ondanks symptomen gewoon doorwerken.......' (!!!!)
'Coronavirus: alarmerend aantal dodelijke slachtoffers mRNA vaccins' (!!!!) (en zie ook de reactie onder dat bericht)
'Coronacrisis: Australische militairen moeten de straten en huizen van o.a. moslims in Sydney controleren' zie wat betreft Australië en de Coronamaatregelen ook:
'Frankrijk en Griekenland voeren vaccinatieplicht in voor verpleegkundigen en verzorgenden' (en zie de links in dat bericht!!)
'COVID-vaccin slachtoffers openen website na censuur van media die imago van vaccinmakers belangrijker achten dan volksgezondheid' (!!!!) (en zie de links in dat bericht!!)
-----------------------------------------------
Over de betrouwbaarheid van de farmaceuten (zie daarvoor ook het eerste blok met links over Pfizer): 'British Medical Journal eist onmiddellijke vrijgeving van alle gegevens over COVID-19 vaccins en onderzoek naar veiligheid'
'COVID-19 vaccin ontwikkeld in Texas: patentvrij voor de wereld' Terwijl de farmaceutische maffia (de grote farmaceuten) weigeren om hun recept gratis door te geven aan arme landen......
'COVID vaccinmakers: hoe betrouwbaar zijn deze farmaceuten'(!!!!)
'COVID-19: internist, cardioloog en epidemioloog wijst op gevaar van vaccins'
'Coronavaccinatie ook als je weet dat de fabrikant ernstige bijwerkingen niet uitsluit?' (!!!!) en zie:
'Coronavaccinatie in vergelijking met het kopen van een huis (waar farmaceuten elke verantwoordelijkheid voor bijwerkingen afwijzen!!)' (en zie de links in dat bericht)
'Coronavirus:
Diederik Gommers (IC arts) zou het mooi vinden dat mensen worden
ingeënt met het AZ vaccin nadat ze tekenden voor het risico dat ze
lopen.....' (!!!!) (en zie de links in dat bericht!!) Een
bericht gebracht nadat bleek dat veel mensen slecht reageerden op het
AstraZeneca vaccin.... Eenzelfde eis overigens die de grote farmaceuten
aan de landen hebben gesteld waar hun vaccin werd verspreid, dus geen
gerechtelijke vervolging voor ernstige bijwerkingen op de korte,
middellange en lange termijn, kortom zelfs de makers vertrouwen hun
eigen vaccin niet......
'COVID-vaccin slachtoffers openen website na censuur van media die imago van vaccinmakers belangrijker achten dan volksgezondheid' (en zie de links in dat bericht!!) (!!!!)
'Coronabeslommeringen: AstraZeneca vaccin onder vuur >> universiteit van Oxford start opnieuw met angstzaaien voor het virus' (universiteit van Oxford heeft het vaccin gemaakt met AstraZeneca.......)
'Astra Zeneca volkomen veilig, uh nee niet voor ouderen, uh oh toch veilig voor ouderen, uh maar niet voor vrouwen onder de 60, uh ho even: toch wel, uhhhh.....'
-----------------------------------------
Wat betreft de betrouwbaarheid van de PCR test zie: PCR test in VS meer en meer onder vuur, ook de FDA en CDC willen ermee stoppen' en zie ook de volgende 5 links naar berichten over de PCR test en de waardeloze test op antilichamen:
'Coronavirus: PCR-test kan niet aantonen dat het virus aanwezig is in het lichaam, beweringen dat dit wel zo is zijn gelogen' In dat bericht zie je een groot aantal links naar oudere artikelen over het Coronavirus)
'Uitvinder PCR test (algemeen gebruikt voor COVID-19): onbetrouwbaar als medische test' Hier een video over deze zaak met de uitvinder en de daarvoor met de Nobelprijs beloonde Kary Mullis:
'Coronavirus, PCR test: leugens en onzin' Een artikel van Mario Ortiz Buijsse.
Ook de test op antilichamen klopt
niet, dit is een test voor mensen die het virus onder de leden hebben
gehad (!!!!):
'COVID-19
testen op antilichamen van o.a. Roche, Quest en Abbott zijn
waardeloos'

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
De volgende reactie ontving ik per mail, deze is afkomstig van Engelbert Valstar en belangrijk genoeg om te plaatsen:
BeantwoordenVerwijderenHet betreft in vitro-onderzoek van slechte kwaliteit,. waarin een aanwijzing zou zijn gevonden dat DNA gerelateerd aan het Covid-19 virus in het lever -DNA zou zijn ingebouwd. Er zijn 2 problemen: het Covid-19 virus (bestaande uit eiwitten, enzymen en RNA) maakt het enzym reversetranscriptase niet (noodzakelijk om dergelijk DNA te maken) en het Pfizer-vaccin is heel iets anders : dit bestaat uit een stukje RNA met daaraan vast synthetisch RNA ; over dit vaccin gaat het hele onderzoek niet! Als biochemicus zeg ik derhalve dat dit verhaal van een linkedin-account niet klopt. Er hebben zeer veel mensen aan het Pfizer-vaccin hun leven te danken en die bewering is op zeer goed onderzoek in vivo, bij zeer grote aantallen, gebaseerd.